
Biocomplexity, Spatial Scale, and Fragmentation: Implications for Arid and Semi-arid Ecosystems (SCALE)

Proposal
Download a printer-friendly pdf version of
the proposal.
(Must have Adobe Acrobat Reader to view. Click here
to download.)
Biocomplexity,
Spatial Scale and Fragmentation:
Implications for Arid and Semi-arid Ecosystems (SCALE)*
Principal Investigator and Co-PI's: J. Ellis1, N.T. Hobbs1, R. Behnke2, P. Thornton3, R. Boone1. Scientific Investigators: M. Stafford Smith4, A. Ash4, M. Coughenour1, K. Galvin1, J. Gross4, C. Kerven2, K. Price5, R. Reid3, A. Seidl6, C. Togtohyn1. Collaborators: I. Alimaev7, Chen Zhuzong8, J. Dodd9, J. Ludwig4, M. Peel10, M. Rainy3, J. Reardon-Anderson11, D. Sneath12.
*SCALE = Scale and Complexity in Arid Land Ecosystems. 1. Natural Resource Ecology Laboratory (NREL), Colorado State University; 2. Macaulay Land Use Research Institute (MLURI) UK; 3. International Livestock Research Institute, (ILRI) Nairobi, Kenya; 4. Commonwealth Scientific and Industrial Research Organization (CSIRO), Australia; 5. Kansas Applied Remote Sensing Program (KARS), Kansas University; 6. Agricultural and Resource Economics Dept., Colorado State University; 7. Institute of Pasture and Fodder, Al Maty, Kazakstan; 8. Institute of Botany, Beijing, China; 9. Cameron University; 10. Agricultural Research Council, South Africa; 11. Asian Studies Program, Georgetown University; 12. Faculty of Archaeology and Anthropology, Cambridge University, UK.
Preface
Overview
Conceptual Framework
Research Goal and Global Objectives
Research Design
Specific Research Objectives and Methods
Results and Implications
Literature Cited
PROJECT DESCRIPTION:
PREFACE: This proposal is a re-submission.
It was previously submitted to the 2000 Biocomplexity competition. It
was not selected for funding in 2000, but did receive ratings of competitive
and highly competitive and was recommended by the Biocomplexity Panel
to be revised in response to comments provided by the panel and reviewers.
The main revisions the panel suggested, and our responses to them, are
listed here. Suggestions: 1) More methodological detail; We have included
more detail in the Research Design, Research Objectives and Methods Sections.
2) Improved linkages between the socio-economic and ecological portions
of the study; See section on this topic. 3) Explanation of how the results
of the study will move us to a higher level of understanding; see Results
and Implications. 4) Better definitions of roles for the large number
of investigators; see Research Design. Some additional points were raised
in the Biocomplexity Advisory Panel Summary. These included: 5) Remote
sensing experiments are not described sufficiently; more detail on remote
sensing techniques is included in research objective 2. 6) Need a better
description of comparability of sites and data collection techniques;
see Research Design section and table 2. 7) Animal movement is not explicitly
addressed; we now address this directly in several places. 8) It is not
clear how SAVANNA model use differs from current work; the model has previously
been adapted to some of our research sites, but never before used to investigate
ecosystem complexity, fragmentation or time-space interactions.
Overview
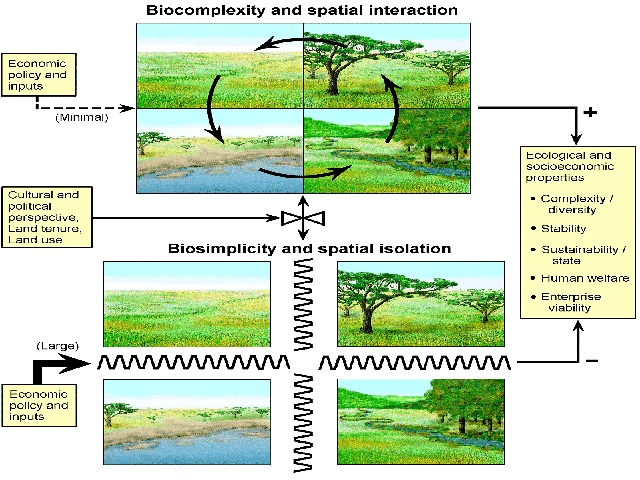
Biological complexity in arid and semi-arid lands (ASALs) arises from
spatially-linked ecological states and processes. Herbivores, humans and
other agents integrate distinct spatial units into complex ecosystems
by moving among and exploiting these units. Spatial complexity plays a
central role in the structure and function of grazed ASAL ecosystems,
but modern human land use tends to deplete spatial biocomplexity through
ecosystem fragmentation. Ecosystems are simplified by breaking up interdependent
spatial units into separate entities, compartmentalizing ecosystems into
isolated sub-units (Fig.1). The result is a reduction in the scale over
which complex interactions among environment, large herbivores and human
management take place. Ecosystem fragmentation and the reduction of biocomplexity
interferes with ecosystem function and reduces system capacity to support
ecological communities, social structures and economic activities. As
a result, many of the world's ASAL ecosystems are dysfunctional to varying
degrees. Dimensions of dysfunction vary from place to place, but include:
increasing conflicts between wildlife and humans (19); wide-spread rangeland
degradation in East Asia (82); increasing levels of poverty among pastoral
people in Africa (55,88); the decline of rural livelihoods in the rangelands
of Australia and the western US (72,84); wholesale collapse of grazed
systems in Central Asia (30,5, 51,52); and global-scale outbreaks of livestock
diseases (‘mad-cow,' foot and mouth disease) in confined industrial
livestock enterprises. Our team's global research experiences suggest
world-wide fragmentation of biocomplexity in ASAL grazinglands caused
by a complex, but discordant, set of interactions involving ecosystem
spatial properties, economic concepts, and legal-political constraints
on land tenure and land use. Improvement of the situation will require,
among other things, a much better scientific understanding of complex
interactions among ecological, political and economic systems.
{kind=link}
Conceptual
Framework
Biological complexity, spatial scale and connectivity in arid and semi-arid
ecosystems
Arid and semi-arid lands (ASALs) are not generally thought of as particularly
diverse or complex. But complexity arises across gradients of climate,
soils, landscape and disturbance (18). Thus complexity is scale-related,
but depends on spatial linkages among ecological states and processes;
i.e., individual sites may not support much complexity, but when linked
together across gradients, they form complex ecosystem states and allow
for complex processes. Said another way, ASAL ecosystems might be thought
of as simple, rather than complex, except for the fact of spatial linkages
and interaction among spatial units. Spatial complexity is crucial in
ASAL ecosystems, which tend to be both spatially extensive and temporally
variable. Spatial scale and access to biocomplexity offsets the destabilizing
effects of temporal variability (8,64,24,25).
Central Theme: ASAL ecosystems function as complex, integrated systems by virtue of connectivity among sets of less complex units. Complexity offsets the effects of temporal variability.
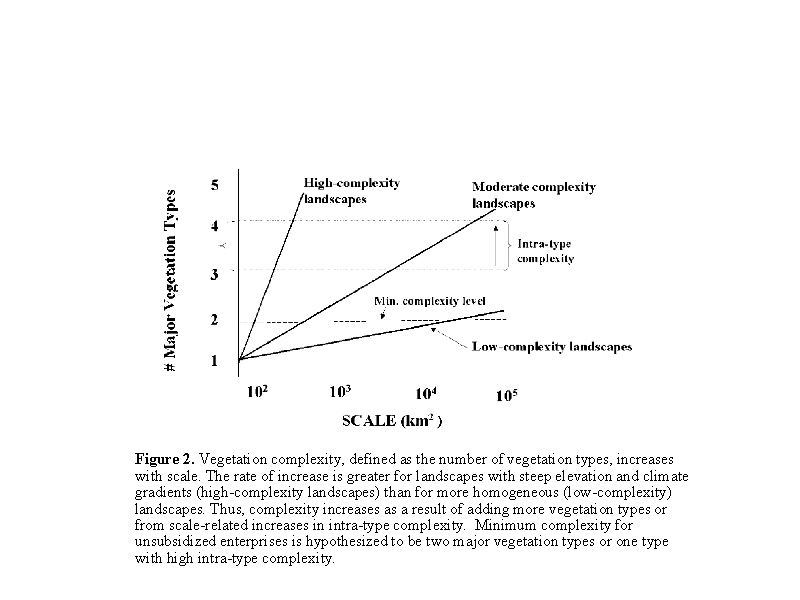
In complex ecosystems, large herbivores shift their ranges from one eco-zone to another through seasonal and interannual cycles. Vegetation complexity creates the opportunity for selectivity and alternatives that reduce the effective amplitude of seasonal and annual variation in food abundance and quality (23,27). In this way, vegetation complexity stabilizes individual condition and population performance by dampening temporal variability in food supplies. Large herbivores require access to a complex set of vegetation communities to maintain animal condition, productivity, and population stability, especially under conditions of high climatic variability (23,24,14,27,3,42,43). We hypothesize that: 1) ASAL ecosystems have an optimal spatial domain for herbivore-based exploitation; and 2) the appropriate domain is under the strong influence of temporal variability and patterns of vegetation complexity. We expect that the minimum level of vegetation complexity for unsubsidized large herbivore exploitation systems will incorporate at least two (and often more) distinct vegetation communities, each having different forage production patterns and forage quality attributes. This translates into wet-dry season ranges in tropical regions or summer-winter ranges in temperate zones. Low-complexity environments require large-scale exploitation strategies (Fig. 2) to access the minimum level of biocomplexity (45). High vegetation complexity (represented by two or more vegetation types in Fig. 2) provides more selectivity options so that minimum complexity is attained at smaller scales (42).
{kind=link}
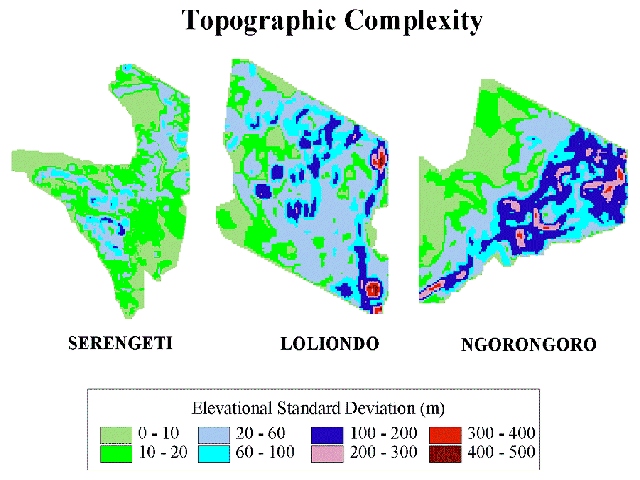
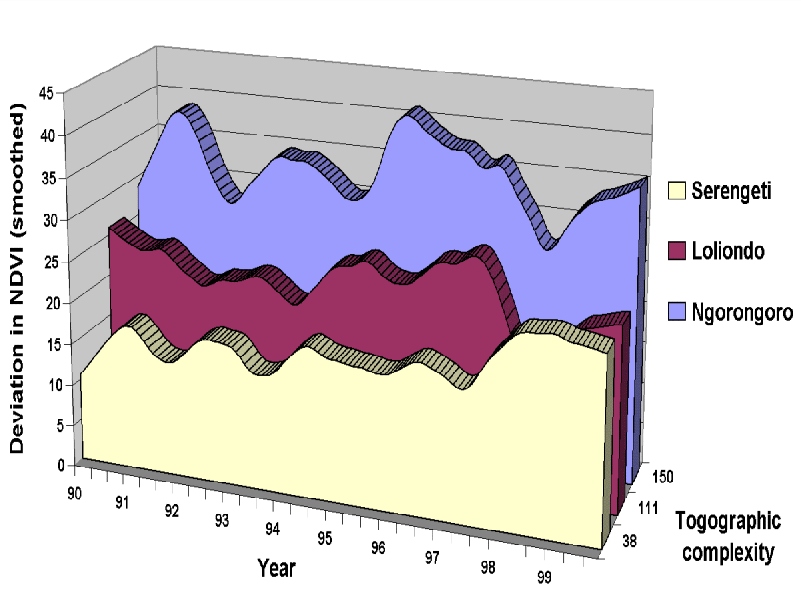
The ecological dynamics behind these concepts are illustrated in Figures 3a and 3b with data from our research in northern Tanzania (6,55). Topographic complexity (calculated as a moving window standard deviation of gridded elevation) is assumed for illustrative purposes as a surrogate for vegetation complexity. Complexity arises in these systems mainly from elevation and rainfall gradients. These three adjacent ecosystems (Fig. 3a) range from moderate complexity, in the shallow-gradient Serengeti National Park, to high complexity in conjunction with steep gradients in Ngorongoro Conservation Area. Loliondo Game Control Area gradients are intermediate, but mild topographic relief causes recurring vegetation complexity there. Vegetation spatio-temporal dynamics (standard deviation of NDVI units) (Fig. 3b) and herbivore movement patterns reflect vegetation spatial complexity. Shallow-gradient systems support lower vegetation complexity (Serengeti Fig. 3a), thus at any particular time, spatial variation in vegetation is likely to be relatively low (Serengeti Fig. 3b). Steep-gradient systems support greater vegetation complexity (Ngorongoro 3a) with greater spatial variation at any particular time (Fig. 3b). Herbivores respond to these diverse patterns of complexity with different movement patterns. For shallow-gradient systems, both wild and domestic herbivores tend to make long distance, semi-nomadic movements (e.g., Serengeti wildebeest and Turkana pastoralists in northern Kenya (23,50,51,60,70). In steep-gradient systems, herbivores and herdsmen undertake seasonal migrations between highlands and lowlands (e.g., elk herds in the western US and Maasai livestock in Ngorongoro (70,43)). Where gradients and complexity are intermediate, as in Loliondo, pastoral livestock utilize a large home range differentiated into dry and wet season grazing zones (55). The same is true for resident wildlife in the Maasai Mara Game Reserve, where seasonal movements within home ranges pulsate around ‘hot spots' expanding during the dry season and collapsing in the wet season (75).
Central
Theme: Connectivity among landscape units is established through movements
undertaken by herbivores, humans and other agents in the process of exploiting
these differing units. Movement among landscape units is an important
process organizing ecosystem complexity, creating the opportunity for
selectivity, and providing alternatives that reduce the amplitude of seasonal
and annual variation in forage abundance and quality.
{kind=link}
{kind=link}
Land use, land
tenure and fragmentation of ASAL ecosystems
Although movement-mediated connectivity is a crucial attribute of ASAL
ecosystems, human land use and land tenure systems tend to fragment ASAL
ecosystems into disconnected parcels (Fig. 1). Fragmentation occurs with
the imposition of a land tenure system, usually to facilitate protection
or usurpation of some key portion of the ecosystem, to implement private
property rights, promote economic intensification, enforce sedentarization
of nomads, or to facilitate other policies or political agendas (30,34,67,84,5).
Four idealized property systems (Table 1) provide the theoretical justification
for different types of land tenure regimes. These idealized systems are
distinguished by characteristic property-owning units and by the distinctive
mechanisms intended to control rates of resource exploitation for each
property type.
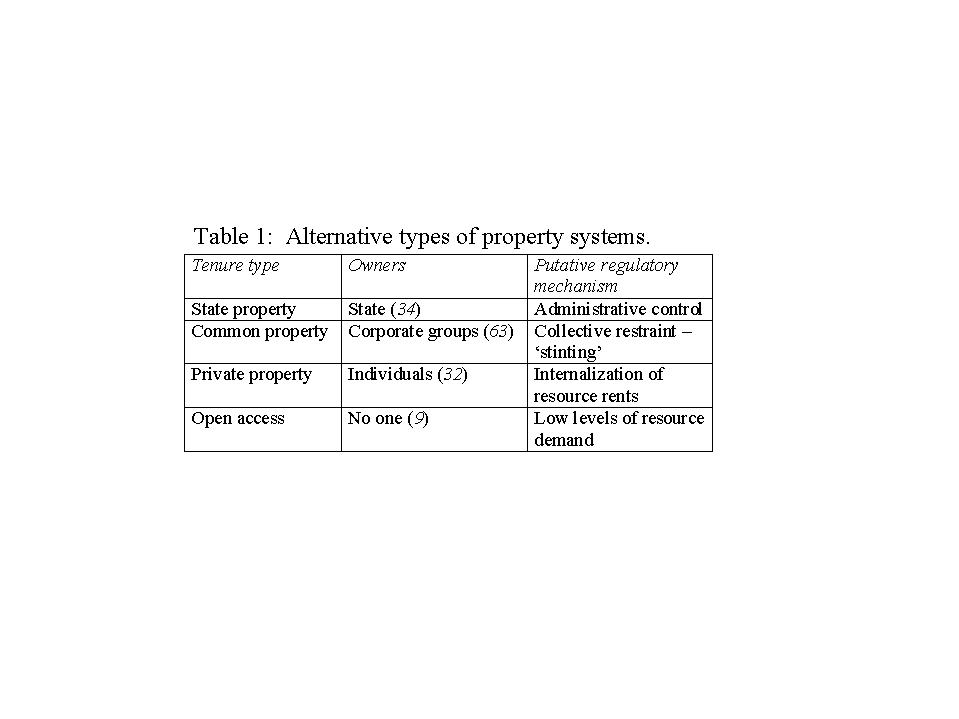{kind=link}
These theoretical forms of land tenure have been used to understand existing property rights regimes, and - more polemically - to create these systems by influencing policy. Each property type has been appropriated by one of the grand theories of political economy including capitalism, communism, and Euro-American notions of primitive political systems. For our purposes, it is noteworthy that fragmentation, justified in different ways in different political systems, is a near-universal feature of modern land tenure systems. Today's dominant concepts of land tenure developed and flourished in the relatively mesic environments of western Europe and eastern North America. The transfer of these mesic tenure systems to arid and semi-arid ecosystems has caused ecological damage and economic disruption (4,24,90,45). Although benefits, such as ease of management and security of investment, may arise from fragmentation. other results are far from beneficial (Fig.1).
Central
Theme: Political and economic imperatives favor fragmentation and the
removal of connectivity of ASAL rangelands. Although benefits, such as
ease of management and security of investment, may arise from fragmentation,
it compromises ecosystem function and the viability of grazing systems
by restricting movements and reducing access to ecosystem complexity.
Economic dimensions
of ecosystem fragmentation
Neo-classical economic perspectives routinely under-value ecosystem natural
capital resources and assume these can be perfectly substituted by economic
inputs (71). Thus, fragmentation and loss of access to biocomplexity are
not perceived as negative aspects of development or land use, but rather
as necessary steps toward intensification and economic growth. Economic
inputs may be rewarded by higher regional carrying capacity and productivity
per unit area, but in the past, the value of biocomplexity has not been
costed properly, only the economic side of the equation is considered;
the ecological side and it's value are ignored. However, ecosystem scientists
and ecological-economic practitioners understand that complex systems
are self-sustaining, whereas simplified (fragmented) ecosystems often
require capital inputs, subsidies and/or management to be sustainable
(Fig.1). While ASAL ecosystem fragmentation is often justified as a means
of economic intensification in the neo-classical framework, in fact, it
costs money (fodder, infrastructure, etc.) to replace the access to natural
capital lost through fragmentation (71). Land use patterns, driven by
economic or political agendas, are unlikely to be perfectly superimposed
on spatial complexity patterns. Where land tenure dictates a small-scale
pattern of exploitation, economic inputs are needed to compensate for
the natural capitol lost to fragmentation. We hypothesize that inputs
per unit area increase exponentially with fragmentation and decreasing
scale (Fig. 4). Alternately, scale expansion through consolidation (Fig.2
) adds greater complexity to the grazing orbit, reducing economic inputs
until at some larger scale, the minimum level of complexity for unsubsidized
exploitation is reached, and economic inputs approach zero (Fig. 4).
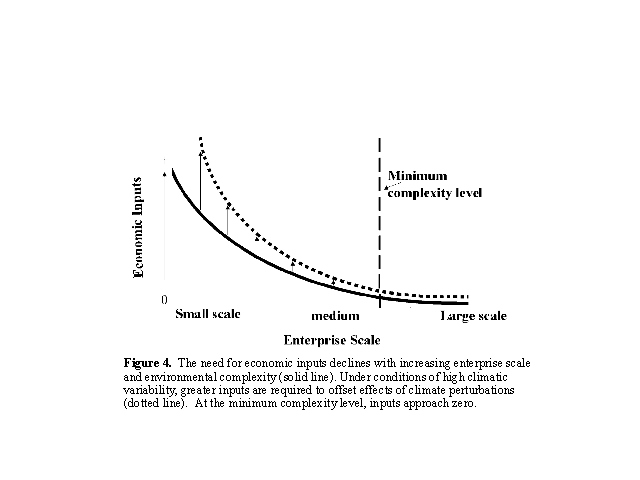{kind=link}
A critical issue is to understand the trade-offs between loss of access to biocomplexity and the benefits of intensified land use, given different forms of economic substitution and a proper ecological economic accounting of natural ecosystem values. To the best of our knowledge, this sort of economic assessment has not been conducted, although many of the building blocks to permit such an analysis are in place.
Central
Theme: Human land tenure or land use patterns, dictated by political or
economic imperatives, are seldom superimposed on ecosystem spatial complexity
patterns. Where land tenure dictates a sub-optimal scale of exploitation,
economic inputs are required to compensate for the natural capital lost
to fragmentation. Benefits derived from economic subsidies may or may
not compensate for the loss of biological complexity.
Research Goal and Global Objectives
We argue that vegetation complexity and spatial scale are crucial but
diminished components of ecosystem function. Our goal is to demonstrate
the importance of complexity and the costs of fragmentation at sites around
the world, by linking ecological and socio-economic research, and in the
process, create an international network of scientists addressing these
issues. Our global objectives are:
1. Develop a framework for analyzing and describing ecosystem spatial
complexity and its role in grazed ecosystem function and sustainability,
including the movement-mediated responses of herbivores to complexity
and fragmentation.
2. Determine the effects of real fragmentation experiments on herbivores,
ecosystems, enterprises and people (Fig.1), and use model-simulated fragmentation/consolidation
experiments to identify options for ecological and economic sustainability.
3. Characterize patterns of ecosystem fragmentation as they exist under
different environmental, political and economic systems; investigate how
and if ecological and political-economic factors interact to control the
evolution of land use systems.
4. Create a method and modeling approach for assessing the value of natural
capitol in complex grazed ecosystems, the costs of complexity loss due
to fragmentation, and the trade-offs between economic inputs and ecological
complexity.
5. Coordinate these analyses in an integrated assessment of complexity
and fragmentation.
Research Design
These objectives would be difficult to achieve under most circumstances.
These problems operate over large spatial scales, long time frames and
involve a variety of disciplines. But a set of natural experiments in
ecosystem fragmentation has been set up (inadvertently) around the world,
allowing us to address these relatively intractable problems. Members
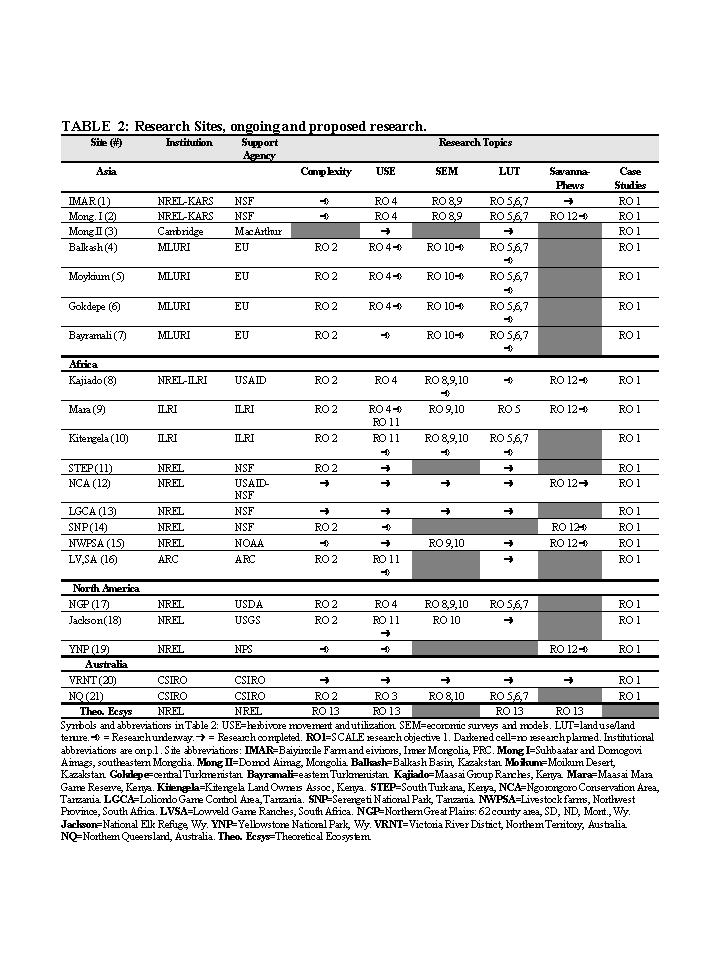
of our team are now working in, or have completed research in 21 ASAL
ecosystems in Asia, Africa, Australia and North America (Table 2). Sixteen
of these ecosystems have undergone fragmentation of one sort or another;
five are more or less intact. These sites/regions form the universe for
our proposed research. This situation presents an unusual opportunity:
a project of this breadth is feasible only because each of the 21 projects
is either partially supported and underway, or recently completed. We
request funds from NSF-Biocomplexity to conduct new research on complexity
and fragmentation at these sites and to integrate and synthesize the results
already obtained by creating a global linkage among scientists; in effect,
merging these projects into an integrated international research program.
Project sites, institutions, funders and a listing of completed, ongoing
and proposed research is presented in Table 2.
Three different types of research are proposed: case study syntheses,
field-based investigations, and model experiments and analyses. (1) Case
studies (RO1) will synthesize information already collected for each site;
however, data and qualitative information will be reviewed and re-analyzed
from a complexity/fragmentation perspective. These 21 case studies will
be completed, presented and prepared for publication in year 1, as the
first activity of the project. (2) Field-based investigations of ecological
complexity, herbivore movements, economic status, and land use/land tenure
patterns are currently (or have been) being conducted at 11 sites (Table
2). New research on these topics is proposed for these 11, plus 4 other
sites. Data collection is complete or available from other sources, for
six sites (3,11,12,13,19,20). (3) Model experiments using SAVANNA-PHEWS,
a linked ecological/socio-economic model, are proposed for seven sites
(1,2,8,9,12,14,15). Application of SAVANNA alone is planned for Sites
19 and 20. An ecological-economic trade-offs model (to be developed in
this project, see RO 10), will be applied at several sites. Finally, a
SAVANNA-PHEWS theoretical version will also be used to address theoretical
complexity-related questions (RO 13).
The project's schedule will focus on the case study synthesis as our first
activity. Field and modeling studies will be the major emphasis in years
two, three and four. Year five will include a heavy emphasis on outreach
as discussed in the next section. We will hold three project workshops
in years one, three and five. The purpose of the first is discussed above.
The year three workshop will provide a forum for discussion of ongoing
field and modeling progress, mid-project synthesis and redirection of
some studies. The year five workshop will be an outreach activity, aimed
at policy and management agencies and institutions.
{kind=link}
Research, education
and outreach
This research project includes graduate students, undergraduate students
and post-doctoral researchers. Research results will be integrated with
educational activities at our five universities. We will conduct outreach
activities to inform policy and management personnel representing international
conservation and development agencies and regional and national agencies,
in each of the nine countries we represent.
Integration
of ecological, social and economic analyses
One of the recommendations made to us by the 2000 biocomplexity panel
was to demonstrate better linkages between the socio-economic and ecological
portions of the study. Sixteen of the site studies in Table 2 involve
integrated ecological and socio-economic investigations; others involve
wildlife populations. The sixteen integrated projects link ecological
and socio-economic components ‘horizontally' in Table 2; all studies
are conducted at the same site, at the same time, using the same informants.
Results are integrated across components to evaluate quantitative relationships.
We will continue to link socio-economic and ecological pattens and interactions
in this way, through bi-variate and multi-variate analyses. In addition,
we integrate through whole-system analyses, (15), most often through integrative
modeling (6). The PHEWS economic model (87) was developed specifically
for linking economic dynamics with SAVANNA ecological dynamics. Whole-system
model (SAVANNA-PHEWS) integration will be conducted at seven sites. Our
proposed economic-ecological trade-offs model will provide comparable
integration a other sites. Promoting site-level integration will be the
responsibility of the coordinating investigator for each site. These are
Ellis (1,2,8,11,17); Sneath (3); Behnke (4,5); Kerven 6,7); Reid (9,10);
Galvin (12,13,15), Coughenour (14,19); Peel (16); Hobbs (18); Ludwig (20);
and Ash (21). Another type of integration is cross-site comparison one
of the great advantages of working simultaneously at several sites. These
will involve whole-system modeling, and analyses of cross-site patterns
for individual research topics. For example, fragmentation takes different
forms in different places. These include breaking up a complex ecosystem
into relatively small properties (Sites 1,3,4,5,8,15,17), lopping off
large critical portions of complex systems (Sites 12,18,19) or excision
of key resources (Sites 1,4,6,7,8,9) Comparing these cross-site patterns
of fragmentation, their origins and implications will provide a clear
understanding of fragmentation as a general phenomenon.
There are gradients within study regions which lend themselves to gradient
analyses. Site 8 for example, has properties ranging from a few hectares
to a few thousand square kilometers. Another form of cross-site comparisons
involves cross-boundary comparisons where the ecosystem is interrupted
by a political boundary, sometimes causing very different dynamics (Sites
1 and 2; 9 and 14; 12 and 13; and 15 contain both private and communal
farms). Coordinating, conceptualizing and facilitating these topical comparisons
will be the responsibility of the PI for each of the seven research topics.
They are: Complexity (Ellis & Boone); Herbivore movements (Hobbs);
Land use/tenure (Behnke); Economic surveys and models (Thornton); SAVANNA-PHEWS
applications (Boone,Thornton).
Specific
Research Objectives and Methods
Specific objectives flow from the global objectives above. We propose
13 research objectives (8 field-based, 5 modeling), many of which will
be applied across all sites. For example, RO 2, complexity analysis, will
be replicated at all sites. ROs 5-8, on land use and economic patterns,
will be applied across all sites. However, research on herbivore movements
varies among sites. Integrated or independent SAVANNA- PHEWS assessments
(ROs 9,12) will take place at thirteen sites. Neither model has been used
before to simulate complexity, fragmentation or time-space interactions.
Research Objective
1: Case Study Synthesis and Comparisons (All scientists and collaborators).
Objective: Develop a state-of-knowledge publication on complexity,
scale and fragmentation. Methods: Scientists involved in this proposal
have, in some cases, years of research experience at their sites. We are
aware of the importance of vegetation complexity, scale and fragmentation
effects. However, with few exceptions, we have not had the opportunity
to investigate these issues directly. We propose to review and re-analyze
existing data and qualitative information gathered at each site, from
the perspective set out in this proposal. The lead scientist for each
site (see Role of Investigators) will organize the site-level synthesis.
Results will be presented at a workshop late in year one, and each synthesis
paper will be prepared in the form of a manuscript, ready for publication.
The PI and co-PI's will develop a cross-site synthesis based on these
individual site reports. This cross-site comparison will establish general
patterns among biocomplexity, fragmentation, economics and symptoms of
ecosystem dysfunction, and will allow us to address synthetic questions
and hypotheses arising from the analysis. The individual site papers and
the cross-site synthesis will be edited and published in book form. We
expect that the book will raise scientific and managerial interest in
the topic and will modify to some extent (but not greatly) the research
plan presented here.
Research Objective 2: Complexity Framework and Analysis (Ellis,
Boone, Price, Reid). Objectives: Develop a framework for complexity
analysis, apply to all sites; determine herbivore access to complexity
for fragmented and un-fragmented grazing orbits (in conjunction with RO
4). Methods: Vegetation complexity will be determined for all sites/regions.
Complexity will be measured in several different ways: simply totaling
the number of distinct vegetation communities per unit area: development
of complexity indices, based on existing vegetation maps (McGarigal and
Marks 1995): from the standard deviation of 1 km NDVI values (Fig 3b):
and the mean of standard deviations of changes in NDVI values for each
pixel, over time. Although vegetation complexity is the primary variable
of interest, broader aspects of landscape complexity will also be investigated.
We will integrate maps of vegetation with elevation and topography, soils,
climate, and land use (where available) through GIS processing. Complexity
will be analyzed at several spatial scales. The smallest scale for the
NDVI data will be 25 km2. Spatial analyses will be scaled up in harmony
with the amplitude of climate variability (seasonal, annual, multi-annual
and long term trends). Alternative measures of complexity will be spatially
correlated to known large-scale patterns of herbivore movements (Sites
5,9,12,13,14,18) and against new data (RO 4), to determine which measures
of complexity provide the best spatial fit to herbivore movements. This
should reveal herbivore responses to vegetation complexity. Differences
in movement between intact and fragmented systems should indicate the
of impact of complexity loss.
NDVI data will be derived from a 1 km AVHRR 10-day Maximum NDVI Composite
(MNC) data set, developed for all 21 research sites, by the Kansas Applied
Remote Sensing (KARS) Program, Kansas University. MNC data sets for most
of 1992-1996 are available at EROS Data Center (EDC). We will develop
additional data sets for 1996-2002, with due consideration for problems
with NOAA 11, 13 and 14. The MNC will be created with standard methods
developed (Eidenshink and Faundeen 1994). Radiometric calibration will
be performed as described by Teillet and Holben (1994). NDVI computation
will use standard formulations (Rouse et al. 1973) and be re-scaled. Imagery
will be geo-referenced using control points from EDC MNC data sets. NDVI
composites will be created on a pixel-by-pixel basis by selecting the
highest NDVI value within a 10-day period (Holben 1986). The MNC will
be corrected for atmospheric attenuation due to Rayleigh scattering and
ozone and to increase the probability of selecting pixels with higher
satellite zenith angles (Chilar and Huang 1994).
Research Objective
3: Herbivore Selection at the Paddock Scale (Ash, Gross).
Objective: Determine the effects of pasture size on animal diet
quality and performance.
Methods: In many tropical ecosystems (i.e., Site 21) annual primary
production may be high, but dietary protein is usually below maintenance
level for much of the year and animal production is limited by diet quality
rather than intake level. The ability of animals to maximize diet protein
is critical to condition and secondary production. We predict that at
similar stocking levels, animal performance is better in large paddocks
than small ones due to greater vegetation complexity, allowing greater
selectivity and improved diet quality. We will investigate this hypothesis
by examining seasonal diet quality of cattle as a function of paddock
size, productivity, and vegetation complexity. Fifty paddocks will be
used, ranging in size from 500 to 5000 ha, stratified across more fertile,
basalt-based soils and nutrient-poor red clay soils that characterize
dry tropical savanna pastures in northern Queensland, Australia (1). We
will estimate cattle diet composition, protein and digestibility in each
paddock, six times per year from the near infrared reflectance signature
of fecal samples (56,57). Animal weight gains and losses will be assessed
periodically with the paddock owner. For each paddock, vegetation complexity
will be mapped from Landsat TM or MODIS imagery, and validated from an
extensive CSIRO data base and from additional sampling. By sampling many
paddocks varying along gradients of complexity, fertility and size, we
will obtain data necessary to quantify the effects of area, vegetation
complexity, and soil type on diet quality and animal performance. This
research addresses herbivore selection at intermediate and large time-space
scales (within and between seasons) and addresses global objectives 1
and 2.
Research Objective 4: Herbivore Movements in Fragmented vs Intact Ecosystems (Ellis, Reid, Behnke).Objective: Determine effects of fragmentation on herbivore access to ecosystem complexity. Methods: This field study will evaluate movements of herbivores and their access to vegetation complexity on properties or grazing areas of various sizes, from a few hectares to several thousand square kilometers. Movements over several small to medium-sized properties will be determined at Sites 1,8,10, 21. Movements by pastoral herders covering medium to large areas will be determined at Sites 2,4,5,6,7,9 (already done at 12,13). Extant data on wild herbivore movements are available for Sites 5,9,14,18. Tempo-spatial patterns of vegetation complexity will be obtained from RO 2. Information on livestock herd movements will be obtained at all sites by interviewing herders (55). Interviews with ~50 herders will be conducted at each site. Herders will be asked to recount seasonal herd movements starting in 1992 through 2002; and to also describe movements during exceptional (i.e., drought, etc.) years. Herd destinations will be located on the ground, described and GPS'ed. Herders will be asked to identify kinds and quantities of supplemental feed where applicable. At Sites 1,5 and 8, year-long herd tracking and GPS plotting will also be conducted. At Site 8 we will evaluate the effects of differential vegetation access among properties of different scale, on livestock diet quality by NIRS fecal analysis as described in RO 3. Livestock condition indices will be estimated four times per year at sites where we have on-site personnel (Sites 4,8,9,10). The effect of fragmentation on wild herbivore abundance and diversity will be investigated three African sites (9,10,16) representing different points on the fragmentation continuum. Variables analyzed will include property size, vegetation complexity patterns (RO 2), livestock herd size, livestock condition index, supplemental feed provided, NIRS diet quality, and other factors collected in RO 8.
Research Objective
5: Typology of Actual Land Use Patterns (Behnke, Kerven, Galvin, Reardon-Anderson,
Gross). Objective: Develop a standard format to differentiate and
compare land use patterns and management scales within and across study
sites.
Methods: We will use published and unpublished data to catalogue
both the customary usages and the legal regulations governing land use
at Sites 1,2,4-10,15,17,21. Information on land use patterns and seasonal
livestock movements, will be obtained in conjunction with RO 4, from a
sample of large and small herd owners possessing different kinds of land
entitlements and occupying large to small scale areas. This information
will be used to compare herd owners' resource entitlements to their actual
patterns of access and land use. Based on these accounts, we will construct
land use and ownership matrices that characterize the size and type of
the household or enterprise units, the different kinds of resources controlled
by each, the spatial extent of their entitlements, and any restrictions
on access or use. These matrices will provide a standard format for characterizing
the essential features of tenure systems and, by abstraction, allow comparison
across sites with different legal and cultural traditions. By comparing
data on actual land use with ecologically optimal land use patterns estimated
in RO 2, we will identify, for several sites, the divergence between the
scale at which resources are actually managed in fragmented systems and
the scale at which they should be managed to achieve economic and ecological
objectives. This research addresses global objective 3.
Research Objective 6: Origin, Evolution of Land Tenure Patterns (Behnke, Reardon-Anderson, Galvin). Objective: Analyze the history of land use systems; their causes and effects. Methods: A qualitative assessment of the cultural and legal variables influencing the evolution of pastoral property systems will be conducted at several study sites, where land tenure patterns have been and remain, dynamic and changing (Site 1,2,4,5,6,7,8,10,15,17,21). We will analyze how these land use systems have evolved in response to ecosystem patterns and dynamics, and in response to political and legal pressures. Analysis will focus on the last century, and will forecast the outcomes of current legal, policy, and economic conditions. The historical studies will focus on how pastoral communities appropriate national land policies and manipulate them for their own ends. Specifically, we will examine: 1) land use policies at different historical periods; 2) interpretation of these policies by local land users; 3) material concerns that motivate the interpretive process; 4) the relationship between ideology and observable patterns of land use; and 5) if and how ecological variations over time and space have influenced local conceptions of property rights. These analyses will determine how national policies have interacted with local land use systems and will identify the mechanisms of this interaction. Analysis will be based on: 1) written historical material, including academic studies, legislation, cadastral surveys and court records; 2) interviews with land users and local administrators.
Research Objective
7: Factors Driving Contemporary Trends in Land Use Change (Behnke,
Galvin, Reardon-Anderson, Kerven, Gross). Objective: Investigate
how ecological, political and socio-economic factors interact to influence
individual land use decisions.
Methods: Following the results of RO 6, we will investigate ecological,
political and socio-
economic variables that influence producers' land use decisions, how these
individual decisions are aggregated into new patterns of land use and
whether the current trend is toward fragmentation or consolidation. Work
will focus on regions in which land use systems are currently in rapid
flux: 1) Central Asian rangelands (Sites 4,5,6,7) where the demise of
the Soviet Union is causing the reorganization of land use; 2) South Africa,
(Site 15) where recent changes have altered the political and economic
environments of both commercial and communal pastoralists; and 3) semi-arid
Australia (Site 21) and North America (Site 17) where long term economic
trends have rendered many private farms and ranches unviable. Our ongoing
studies in these regions closely complement these objectives. Data collection
will include factors like: enterprise type and diversity, livestock numbers,
human population densities, socio-economic characteristics, and most particularly,
the constraints and incentives that influence stock managers' decisions
(see RO 8). Information will be collected for 20 households per site.
Information on resource use patterns will coordinated with RO 4. We will
interpret PHEWS assessments (RO 9) to evaluate the relative weights of
these different factors in driving land use change trends.
Research Objective
8: Economic Surveys and Analysis (Thornton, Stafford Smith, Seidl).
Objectives: Gather information on household economic performance
and the economic dimensions of livestock production systems in relation
to scale and resource access.
Methods: Economic data at household and enterprise levels are necessary
to determine relationships among complexity and economic welfare as proposed
in global objective 4. Economic data will be obtained for all project
sites supporting commercial or subsistence livestock enterprises (Sites
1,2,4,5, 6,7,8,9,10,15,17,21). Data will focus on the drivers of household
livelihoods and enterprise economic viability, their spatial resource
access patterns, and their levels of material or financial subsidy. The
general approach to gathering this information is: 1) literature search
of secondary data sources; 2) the design and testing of survey instruments
to elicit information from pastoralists; 3) data collection using the
survey instrument and, where necessary, informal interviews with key informants.
Data analysis and interpretation will be carried out by assembling household
budgets and by constructing decision trees for major pastoralist decisions.
This information is critical in terms of understanding the pastoral systems
under study and the question of how enterprise scale and access to ecological
complexity relates to economic status. In addition, it forms the basis
for research objective 10.
Research Objective 9: PHEWS (Pastoralist Household Economic Welfare Simulator) Model Assessments (Thornton, Stafford Smith, Seidl). Objective: Determine economic-ecological interactions resulting from alternative land use practices. Methods: This research will simulate and predict economic consequences of alternative land tenure/land-use practices, under various social and ecological conditions. Scenario analysis will be carried out to assess possible impacts of infrastructural and policy changes on household and enterprise incomes, particularly with regard to changes in access to vegetation complexity under different management or land tenure regimes. We will use existing household and enterprise-level models to 1) determine the levels of vegetation complexity and environmental resources needed for subsistence with minimal economic inputs, and 2) how inputs increase (if they do) with increasing levels of fragmentation. Existing models to be used at SCALE sites include: 1) a rule-based household food security and cash flow and household decision model (87), embedded in SAVANNA; 2) a multi-objective mixed farm household model for communal and semi-commercial farming systems in southern Africa (Herrero, Thornton and Galvin, in progress); and 3) a herd dynamics and enterprise economics model for commercial farms (Herd-Econ, see 82). The general approach includes a number of steps. First, adaptation of existing models to new case study sites, (partially fed from surveys carried out in RO 10 above). Second, calibration of the models, using existing data sets. Third, development of scenarios to be assessed. Fourth, scenario analysis and assessment of the results.
Research Objective
10: Trade-offs model: development and assessment (Stafford Smith,
Thornton, Seidl). Objective: Determine the integrated trade-offs
between ecosystem fragmentation and external economic subsidies, on enterprise
and regional scale productivity.
Methods: We will develop a ‘trade offs' model focusing on
the effects and human responses to fragmentation in ASAL grazed ecosystems.
Based on the outputs of RO 2, but with an explicit incorporation of the
costs and benefits of spatial resource access and the alternative compensatory
mechanisms for this (see Fig. 4), the model will examine how external
subsidy at the household / enterprise scale substitutes for access to
biocomplexity. These findings will be scaled up to the regional scale
to provide the implications to net regional productivity. The initial
phase of model development will assume that grazing dominates land use,
but final analysis will require addressing product differentiation and
land use substitution at the regional level. The model will be parameterised
for a subset of sites, yet to be determined. It will be tested against
observed strategies in regions that are functioning more or less successfully
at present. For example, in Australia, the test will assess the efficacy
of different adaptive strategies (holding multiple leases, trading between
regions, or taking advantage of subsidies) in regions with different levels
of climatic variability and resource fragmentation (e.g., lease size compared
to biophysical heterogeneity), and differential access to markets and
economic productivity. Comparisons with other global systems will allow
us to identify system-level emergent properties (resistance to stress,
resilience, etc.) in conjunction with the model experiments discussed
under RO 12,13, and will permit assessment of the riskiness of different
trade-offs as responses to reduced access to biocomplexity.
Research Objective 11: Spatial Complexity, Temporal Variability and Population Patterns (Hobbs). Objective: Develop competing models linking animal populations to spatial complexity. Methods: Complexity analysis and model selection will be used to test the effects of vegetation complexity on animal population stability. We proposes that landscape complexity offers alternatives allowing animals to cope with temporal variability in ways not possible in fragmented landscapes. If true, then population variability, driven by climatic variability should be ameliorated by spatial variability in un-fragmented landscapes. We will develop competing models predicting population performance of wild ungulates from data on temporal variability in forage and spatial heterogeneity in habitats. We will use time series of field observations on sex and age composition of populations, rate of increase, and total abundance as dependent variables. Independent variables will include time series data on precipitation and temperature as well as indices of vegetative production based on NDVI. To assess landscape complexity we will use indices of landscape heterogeneity and fragmentation derived from vegetation maps (McGarigal and Marks 1995). We have access to detailed data on ungulate populations, vegetation maps and climate observations in seven different study areas in the western United States including Sites 18,19, and similar data for three sites in east and South Africa (9,10,16). We will also attempt to adapt this approach to assess affects of vegetation complexity on herbivore diversity, for the African sites. These data will provide a basis for model selection. Models will range from purely empirical statistical models with few parameters, to highly mechanistic models with many parameters. We will use likelihood-based techniques and information theoretics to assess the best fit to data, among the competing models. If the best approximating models indicate a dependence of the effects of temporal variability on spatial heterogeneity, then our prediction about the role of landscape complexity in modifying effects of temporal heterogeneity will be supported by the observations.
Research Objective 12: SAVANNA-PHEWS Complexity-Fragmentation Experiments (Boone, Coughenour, Thornton). Objective: Model effects of fragmentation on ecosystems and people. Methods: The SAVANNA-PHEWS integrated assessement system (16,17, 87) was created to simulate coupled ecological and economic dynamics of grazed ASAL ecosystems. The model has been or is being adapted to six sites (1,2,8,9,12,15). All except Site 2 are fragmented. Model experiments will investigate the role of fragmentation and loss of access to ecosystem complexity at each site. We will posit a set of alternative land use practices for each site, based on: 1) ecosystem spatial complexity and temporal dynamics; 2) basic economic characteristics; and 3) current and projected human population densities and demands. The potential for wildlife conservation will be factored in at the relevant sites (8,9,12). Simulated alternative land use practices will include greater levels of fragmentation and reductions in fragmentation. Results will examine effects of alternative land use patterns on herbivore condition and dynamics, economic status of residents, and ecological degradation. These alternative land use scenarios will also be used to examine effects on ecosystem stability (measured as stability of livestock populations) and enterprise sustainability (measured as the level and stability of production and offtake) under a variety of climate regimes.
Research Objective
13: Complexity, and Fragmentation in Theoretical Ecosystems (Boone,
Coughenour, Hobbs, Ellis). Objective: Study general responses of
ASAL ecosystems to fragmentation. Methods: We hypothesize that
the effects of fragmentation and complexity-loss cascade through ecosystems
influencing herbivore dynamics, socio-economic systems and ecosystem properties.
These responses will be explored by applying the SAVANNA-PHEWS model to
a theoretical ecosystem that emulates the main components of an African
ASAL, where we can generate alternative structures and processes to identify
general responses, with confidence intervals on metrics. Monte Carlo simulations
(n determined by power analyses) will be conducted using a series of generated
landscapes, to yield confidence estimates. Experimental variables will
include: alternate patterns of temporal (climatic) variability, varying
patterns of vegetation production and complexity; and constraints on herbivore
movements based on different forms of fragmentation or excision of resources.
•Simulations will be organized into three separate nx3 factorial
analyses, with variability in climate (stable, variable, and highly variable)
as a factor in each analysis (25).
•Complexity will be represented by three alternative types: high,
low and intermediate (see Fig. 3).
•Paddock or range sizes will vary to represent unrestricted, large
restricted and small range sizes. We hypothesize (RO 3) that there will
be a non-linear relationship between range size, animal production and
population growth; i.e., that the carrying capacity increases disproportionally
with paddock scale.
•Fragmentation regimes will include: excision of key resources (i.e.,
swamps, riparian zones) with complete access to remaining portions of
the ecosystem; large but critical portions of the ecosystem lopped off
(winter ranges, dry-season ranges); fragmentation into small impermeable
patches; fragmentation into small isolated patches, but connected by corridors.
In each factorial experiment, we will characterize the patterns in herbivore
populations and impacts, ecosystem properties, and human well-being (e.g.,
cash flows). Also, in analyses of increasing fragmentation, we will explore
dynamic and emergent behavior patterns (e.g., resistance, resilience,
unpredictability) of the modeled ecosystems, asking, for example, if the
modeled system becomes more or less stable with increasing complexity.
We will assess the effects of system complexity on non-linear dynamics
by plotting and sectioning attractor domains.
RESULTS
and IMPLICATIONS
The 2000 biocomplexity panel asked how the results of this study will
"move us to a higher level of understanding." In our view, the
research topics (columns, Table 2) are addressing questions not yet studied
very thoroughly in regard to complexity, fragmentation and time-space
interactions. For example, developing a framework and methodology for
defining the optimal spatial domain of grazed ecosystems and how these
domains change with temporal variability seems a new and exciting challenge
with important scientific and practical implications. Likewise, trying
to develop a quantitative means of valuing complexity and the costs of
fragmentation is again, a new, important and interesting challenge. We
think that success in integrating these research topics will provide a
higher level of understanding of spatial-temporal distribution of complexity;
its importance in grazed systems; why, and how complexity is reduced through
fragmentation, and what this means for ecosystems and economic activities.
These results will, furthermore, raise practical questions about modern
land use philosophies and their application and sustainability for ASAL
ecosystems. These questions have been discussed for over a century (71,86);
we intend bring strong scientific evidence to bear on these issues this
time.
LITERATURE CITED
* indicates publication resulting from prior NSF research support
1. Ahern, C.R, Shields, P.G, Enderlin, N.G, and Baker,
D.E. 1994. The soil fertility of central and north-east Queensland grazing
lands. Information Series A194065: MRC and Queensland Department of Primary
Industries, Brisbane, Australia.
2. Ash, A.J. and D.M. Stafford Smith. 1996. Evaluating stocking rate impacts
in rangelands: animals don't practice what we preach. Rangelands Journal
18:216-243.
3. Behnke, R., I. Scoones, and C. Kerven, eds. 1993. Range Ecology at
Disequilibrium. Overseas Development Institute, London, UK.
4. Behnke, R. 1994. Natural resource management in pastoral Africa. Development
Policy Review 12:5-27.
5. Behnke, R. 1999. Reconfiguring property rights in livestock production
systems. Report to the Overseas Development Institute, (ODI) London.
*6. Boone, R., M.B. Coughenour, K.A. Galvin and J.E. Ellis (accepted).
Addressing Management Questions for Ngorongoro Conservation Area, Tanzania,
Using the Savanna Modeling System. African Journal of Ecology.
7. Breiman, L., Friedman, J., Olshen, R. and Stone, C. 1984. Classification
and regression trees. Belmont, CA: Wadsworth.
8. Caughley, G., N. Sheperd and J. Short, eds. 1987. Kangaroos: Their
Ecology and Management in the Sheep Rangelands of Australia. Cambridge
University Press, N.Y.
9. Christensen, L., R.M. Reich, M.B. Coughenour, J.E. Ellis, and K.P.
Price. In review. Correcting for spatial autocorrelation in detecting
changes in NDVI over time. Ecology.
10. Cihlar, J. and Huang, F. 1994. Effect of atmospheric correction and
viewing angle restriction on AVHRR data composites. Canadian Journal of
Remote Sensing,20(2):132-137.
11. Ciriacy-Wanthrup, S.V. and R.C. Bishop. 1975. Common property as a
concept in natural resource policy. Natural Resources Journal 15:713-27.
12. Cliff, A.D. and J.K. Ord. 1981. Spatial processes: models and applications.
Pion Limited, London, UK.
13. Cook, R.D. and S. Weisberg. 1990. Confidence curves in nonlinear regression.
Journal of the American Statistical Association 85:544-551.
*14. Coughenour, M.B. 1991. Spatial components of plant herbivore interactions
in pastoral, ranching and native ungulate ecosystems. J. Range Mgmt. 44(6):530-542.
*15. Coughenour, M.B., J.E. Ellis, D.M. Swift, D.L. Coppock, K. Galvin,
J.T. McCabe, and T.C. Hart. 1985. Patterns of energy extraction and utilization
in a pastoral ecosystem. Science 230:619-625.
*16. Coughenour, M.B. 1992. Spatial modeling and landscape characterization
of an African pastoral ecosystem: a prototype model and its potential
use for monitoring droughts. In: D.H. McKenzie, D.E. Hyatt, and V.J. McDonald,
eds. Ecological Indicators, Vol. I. Elsevier Applied Science, London and
New York, pp.787-810.
*17. Coughenour, M.B. 1993. The Savanna landscape model - Documentation
and Users Guide. Natural Resource Ecology Laboratory, Colorado State University,
Fort Collins CO.
*18. Coughenour, M.B. and J.E. Ellis. 1993. Climate and landscape control
of woody vegetation in a dry tropical ecosystem, Turkana District, Kenya.
Journal of Biogeography 20:383-398.
19. Crowe, D. 1995. Status of selected wildlife resources in Botswana
and recommendations for conservation actions. In: The Present Status of
Wildlife and its Future in Botswana.
20. Cox, C. 1998. The Delta method. pp. 1125-1127. In: P. Armitage and
T. Colton (eds.), Encyclopedia of Biostatistics, Vol. 2. John Wiley &
Sons. Chichester, U.K. Proceedings of Symposium. Kalahari Conservation
Society and Chobe Wildlife Trust. Gaberone. 1995. 11-25.
21. Dent, J.B. and M.J. Blackie. 1979. Systems Simulation in Agriculture.
Applied Science Publishers Ltd, London.
22. Eidenshink, J.C., and J.L. Faudeen, 1994. The 1 km AVHRR global land
data set: first stages in implementation. International Journal of Remote
Sensing, 15:3443-3473.
*23. Ellis, J.E., K. Galvin, J.T. McCabe, and D.M. Swift. 1987. Pastoralism
and Drought in Turkana District, Kenya. Report to NORAD, Nairobi. 205
pp.
*24. Ellis, J.E. and D.M. Swift. 1988. Stability of African pastoral ecosystems:
Alternate paradigms and implications for development. J. Range Manage.
41(6):450-459.
*25. Ellis, J.E., M.B. Coughenour, and D.M. Swift. 1993. Climate variability,
ecosystem stability, and the implications for range and livestock development.
In: Range Ecology at Disequilibrium. R. Behnke, I. Scoones, and C. Kerven
eds. ODI, London.
*26. Ellis, J. 1994. Climate Variability and Complex Ecosystem Dynamics:
Implications for Pastoral Development. In: Living with Uncertainty. I.
Scoones (ed.) Intermediate Technology Publications, London.
*27. Ellis, J. and K. Galvin. 1994. Climate patterns and land use practices
in the dry zones of Africa. Bioscience 44(5):340-349.
*28. Ellis, J. 1994. Ecosystem dynamics and economic development of African
rangelands: Theory, ideology, events, and policy. In: Environment and
Agriculture: Rethinking Development Issues for the 21st Century. S. Breth
(ed.). Winrock International. Pp. 174-186.
29. Ellis, J. 1999. Extensive Grazing Systems: Persistence under political
stress and environmental risk. Ruminations: Newsletter of the Global Livestock
Collaborative Research Support Program. Winter 1999. P.10.
30. Ellis, J. and R. Lee. 1999. Ecosystem dynamics and ecological perspectives
on the collapse of the livestock sector in southeastern Kazakstan. Report
to the Overseas Development Institute, (ODI) London.
*31. Ellis, J. and M.B. Coughenour. 1998. The SAVANNA integrated modelling
system. In: Drylands: sustainable use of rangelands in the twenty-first
century. V. Squires and A. Sidahmed, editors. IFAD Technical Reports.
Rome. Pp. 97-106.
*32. Ellis, J. and M. Peel. 1995. Economies of Spatial Scale in Dryland
Ecosystems. Arid Zone Ecology Forum. Kimberely, SA. September.
33. Fahrig, L. 1997. Relative effects of habitat loss and fragmentation
on population extinction. J. Wildlife Management 61(3):603-610.
34. Galaty, J. and D. Johnson (eds.) 1990. The World of Pastoralism: herding
systems in comparative perspective. The Guilford Press. New York.
*35. Galvin, K.A. 1992. Nutritional ecology of pastoralists in dry tropical
Africa. American Journal of Human Biology 4(2):209-221.
36. Gamon, J.A., L. Serrano, and J.S. Surfus. 1997. The photochemical
reflectance index: an optical indicator of photosynthetic radiation use
efficiency across species, functional types, and nutrient levels. Oecologia
112:492-501.
37. Gordon, H.S. 1954. The economic theory of a common-property resource:
the fishery. Journal of Political Economy 62:124-42.
38. Grandin, B.E. 1991. The Maasai: socio-historical context and group
ranches. Maasai herding: an analysis of the livestock production system
of Maasai pastoralists in eastern Kajiado District, Kenya. S. Bekure,
P.N. de Leeuw, B.E. Grandin and P.J.H. Neate. Addis Ababa, Ethiopia, International
Livestock Centre for Africa.
39. Hardin, G. 1968. The tragedy of the commons. Science 162:1243-48.
40. Hart, R.H., J. Bissio, M.J. Smauel, and J.W. Waggoner Jr. 1993. Grazing
systems, pasture size, and cattle grazing behavior, distribution and gains.
Journal of Range Management 46:81-87.
41. Hobbs, N.T. 1996. Modification of ecosystems by ungulates. Journal
of Wildlife Management 60:695-713.
42. Hobbs, N.T. 1999. Responses of large herbivores to spatial heterogeneity
in ecosystems. Pages 97-129 In: H.G. Jung and G.C. Fahey (eds.). Nutritional
Ecology of Herbivores: Proceedings of the Vth International Symposium
on the Nutrition of Herbivores. American Society of Animal Science. Savory
IL.
43. Hobbs, N.T., F. Singer, and G.Wockner. 2001. Assessing Management
Alternatives for Ungulates in the Greater Teton Ecosystem using simulation
modeling. Progress Report to Grand Teton National Park.
44. Holben, B.N. 1986. Characteristics of maximum-value composite images
from temporal AVHRR data: The International Journal of Remote Sensing.
7:1417-1434.
45. Humphrey, C. and D. Sneath. 1999. The End of Nomadism? Society, State
and the Environment in Inner Asia. The White Horse Press. Cambridge.
46. Illius, A.W. 1997. Physiological adaptation in savanna ungulates.
Proceedings of the Nutrition Society 56:1041-1048.
47. Illius, A.W., and I.J. Gordon. 1987. The allometry of food intake
in grazing ruminants. Journal of Animal Ecology 56:989-1000.
48. Illius, A.W. and I.J. Gordon. 1992. Modelling the nutritional ecology
of ungulate herbivores evolution of body size and competitive interactions.
Oecologia 89:428-434.
49. Illius, A. and T.G. O'Connor. 1999. On the relevance of nonequilibrial
concepts to arid and semi-arid grazing systems. Ecol. Applications.
50. Inglis, J.M. 1976. Wet season movements of individual wildebeests
of the Serengeti migratory herd. E. Afr. Wildl. J. 14:17-34
51. Kerven, C., and I. Alimaev. 1999. Shifts in the balance between mobility,
fodder and the market. Report to the Overseas Development Institute, (ODI)
London.
52. Kerven, C. and S. Akiner. In press. From State Farms to Private Flocks:
Prospects for Pastoralism in Kazakstan and Turkmenistan. Curzon Press.
London. (in press)
53. Legendre, P. and M.-J. Fortin. 1989. Spatial pattern and ecological
analysis. Vegetatio 80:107-138.
*54. Lynn, S., J. Ellis, K. Galvin, N. Smith. 1999. Ecological Impacts
on Maasai Pastoralist Land Use Patterns: implications for conservation
policy in multiple use areas. Presented at 1999 Landscape Ecology Symposium,
Aspen Colorado. August.
*55. Lynn, S.L. 2000. Conservation Policy and Local Ecology: Effects on
Maasai land use patterns and human welfare in northern Tanzania. MS thesis.
Colorado State University, Fort Collins. 128 pp.
56. Lyons, R.K. and J.W. Stuth. 1992. Fecal NIRS equations for predicting
diet quality of free-ranging cattle. Journal of Range Management 45:238-244.
57. Lyons, R.K., J.W. Stuth, and J.P. Angerer. 1995. Fecal NIRS equation
field validation. Journal of Range Management 48:380-382.
58. Maddock, L. 1979. The "migration" and grazing succession.
In: Serengeti: Dynamics of an Ecosystem. A.R.E. Sinclaire and M. Norton-Griffiths
(eds.). University of Chicago Press. 46-81.
59. Mayer, D.G. and D.G. Butler. 1993. Statistical validation. Ecological
Modeling 68: 21-32.
*60. McCabe, J.T. 1983. Land use among the pastoral Turkana. Rural Afr.
15-16:109-126.
61. McGarigal, K. and B.J. Marks. 1995. FRAGSTATS: Spatial pattern analysis
program for quantifying landscape structure. U S Forest Service General
Technical Report Pnw 0:1-122.
62. Mearns, R. and J. Swift. 1995. Pasture tenure and management in the
retreat from a centrally-planned economy in Mongolia. II. Proceedings
of the 5th International Rangelands Congress. Society for Range Management.
Denver Co.
63. Milner-Gulland, E., R. Mace and I. Scoones. 1996. A model of household
decisions in Dryland Agropastoral Systems. Agricultural Systems. Vol.
51 (4) Pp. 407-430.
64. Nicholls, A.O. 1989. How to make biological surveys go further with
generalized linear models. Biological Conservation 50:51-75.
65. Nicholls, N. and K.K. Wong. 1990. Dependence of rainfall variability
on mean rainfall, latitude, and the southern oscillation. J. Climate 3:163-170.
66. Pastor, J., and R.J. Naiman. 1992. Selective foraging and ecosystem
processes in boreal forests. American Naturalist 139:691-705.
67. Pennycuick, L. 1975. Movements of the migratory wildebeest population
in the Serengeti area between 1960 and 1973. E. Afr. Wildl. J. 13:65-87.
68. Perkins, J.S. and D.S.G. Thomas. 1993. Spreading deserts or spatially
confined environmental impacts? Land degradation and cattle ranching in
the Kalahari Desert of Botswana. Land Degrad. and Rehab. 4:179-194.
69. Peuelas, J., J.A. Gamon, A.L. Fredeen, J. Merino, and C.B. Field.
1994. Reflectance indices associated with physiological changes in nitrogen-
and water-limited sunflower leaves. Remote Sensing of Environment 48:135-146.
70. Peuelas, J., I. Filella, and J.A. Gamon. 1995. Assessment of photosynthetic
radiation-use efficiency with spectral reflectance. New Phytol. 131:291-296.
71. Potkanski, T. 1994. Pastoral Economy, Property Rights and Traditional
Mutal Assistance Mechanisms among the Ngorongoro and Salei Maaai of Tanzania.
Pastoral Land Tenure Series Monograph 2. International Institute for Environment
and Development. London. 156pp.
72. Powell, J.W. (1878). Report on the Lands of the Arid Region of the
United States. Washington DC. Harvard University Press, 1962. Cambridge
Mass.
73. Prugh, T., R. Constanza, J. Cumberland, H. Daly, R. Goodland, and
R. Norgaard. 1999. Natural Capitol and Human Economic Survival. 2nd Ed.
CRC Press, LLC. Boca Raton Florida.
74. Rathge, R. and P. Highman. 1997. Population change in the Great Plains.
A history of prolonged decline. Rural Development Perspectives, USDA.
75. Reich, R. and R. Davis. 1998. Quantitative spatial analysis. Colorado
State University, Fort Collins, Colorado.
76. Reid, R.S. and Swallow, B.M. 1998. Final Technical Report for IFAD
TAG grant No. 284-ILRI, entitled: An Integrated Approach to the Assessment
of Trypanosomiasis Control Technologies and their Impacts on Agricultural
Production, Human Welfare and Natural Resources in Tsetse-Affected Areas
of Africa, ILRI, Nairobi, Kenya. 328 pp.
77. Reid,R.S. and M. Rainy. Research in progress.
78. Rouse, J.W., R H. Haas, J.A. Schell, and D.W. Deering. 1973. Monitoring
vegetation systems in the Great Plains with Third ERTS. ERTS Symposium,
NASA No. SP-351. Pp. 309-317.
79. Runge, C.F. 1986. Common property externalities: isolation, assurance
and resource depletion in a traditional grazing context. American Journal
of Agricultural Economics 63:595-606.
80. Rykiel, E.J., Jr. 1996. Testing ecological models: the meaning of
validation. Ecological Modeling 90: 229-244.
*81. Shipley, L.A., J.E. Gross, D.E. Spalinger, N.T. Hobbs, and B.A. Wunder.
1994. The scaling of intake rate in mammalian herbivores. American Naturalist
143:1055-1082.
*82. Shipley, L.A., D.E. Spalinger, J.E. Gross, N.T. Hobbs, and B.A. Wunder.
1996. The dynamics and scaling of foraging velocity and encounter rate
in mammalian herbivores. Functional Ecology 10:234-244.
83. Sneath, D. 1998. State policy and pasture degradation in inner Asia.
Science 281:1147-1148.
84. Stafford Smith, D.M., and Foran, B.D. 1992. An approach to assessing
the economic risk of different drought management tactics on a South Australian
pastoral sheep station. Agric. Syst. 39:83-105.
85. Stafford Smith, M., Morton, S. and Ash, A. 1997. On the future of
pastoralism in Australia's rangelands. In: Klomp, N. and I. Lunt (eds.).
Frontiers in Ecology: Building the Links. Elsevier Science Ltd, Oxford,
UK. Pp. 7-16.
86. Starrs, P.F. 1998. Let the Cowboy Ride: Cattle ranching in the American
west. Johns Hopkins University Press. Baltimore.
87. Teillet, P.M. and B.N. Holben. 1994. Toward operational radiometric
calibration of NOAA-AVHRR imagery in the visible and infrared channel.
Canadian Journal of Remote Sensing 20(1):1-10.
88. Thompson, D.M. (Ed.) 1997. Multiple Land Use: The Experience of the
Ngorongoro Conservation Area, Tanzania. IUCN, Gland, Switzerland and Cambridge
UK.
89. Thornton, P., K. Galvin and S. Mbogoh. 2000. Integrated Modeling and
Assessment for Balancing Food Security, Conservation and Ecosystem Integrity
in East Africa. GL-CRSP Final Report on Socio-Economic Modeling Component,
1997-2000. ILRI, Nairobi, 87 pages.
90. Urban, D.L., M.F. Acevedo, and S.L. Garman. 1999. Scaling fine-scale
processes to large-scale patterns using models derived from models: meta-models.
Pages 70-98 In: D.J. Mladenoff and W.L. Baker (eds.). Spatial modeling
of forest landscape change: approaches and applications. Cambridge University
Press. Cambridge, U.K.
91. Vermote, E.F., D. Tanre, J.L. Deuzéé, M. Herman, and
J.J. Morcrette. 1997. Second simulation of the satellite signal in the
solar spectrum, 6S: An overview. IEEE Transactions of Geoscience and Remote
Sensing 35(3):675-686.
92. Webb, W.P. 1931. The Great Plains. Waltham, Mass., Blaisdell Publishing
Co.
93. Williamson, D., J. Williamson and K.T. Ngwamotsoko. 1988. Wildebeest
migration in the Kalahari. Afr. J. Ecol. 26:269-280.
*94. Worden, J. Research in progress.
Page Last Modified: October
13, 2006
Disclaimer
| Equal
Opportunity Statement
Webmaster: jlackett@nrel.colostate.edu
